社会的上下関係を巡る闘争で勝敗を分ける神経回路を発見
理化学研究所(理研)脳神経科学研究センター 意思決定回路動態研究チーム(研究当時)の岡本 仁 チームリーダー(研究当時、現 知覚運動統合機構研究チーム 客員主管研究員)、松股 美穂 研究員(研究当時)らの共同研究グループは、マウスの脳にある「手綱核(たづなかく)-脚間核(きゃくかんかく)神経回路[1]」が社会的闘争で勝敗を分ける重要な役割を担っていることを発見しました。
本研究成果は、全ての動物に共通して見られる社会的上下関係を巡る闘争の制御機構の解明につながるだけでなく、鬱(うつ)やひきこもりなどの精神状態の脳科学的理解や治療法の開発に貢献すると期待されます。
岡本 チームリーダーらはこれまでにゼブラフィッシュを使って、脳の手綱核から脚間核に至る2本の神経回路のどちらが強化されるかによって、社会的上下関係を巡る闘争で、勝者になるか敗者になるかが強く影響を受けることを発見しています。
今回、共同研究グループは、マウスも同じ回路を持ち、ゼブラフィッシュの敗者の回路と相同な回路を人為的に弱めると、マウスが、強い相手に対しても闘いを諦めずに継続して負けなくなること、逆に人為的に回路を強めると簡単に闘いを諦めて負けてしまうことを明らかにし、勝敗に影響する神経回路が哺乳類でも同じように働いていることを証明しました。また、この敗者の回路の下流に位置する正中縫線核(せいちゅうほうせんかく)のセロトニン神経細胞[2]を人為的に活性化することで、一度は敗れたマウスを、勝者として振る舞うように変えられることも示しました。
本研究は、科学雑誌『Current Biology』5月5日号の掲載に先立ちオンライン版(4月9日付:日本時間4月10日)に掲載されました。
背景
全ての脊椎動物は、より広い縄張りや、より良い生殖パートナーなどを巡って同種同士で闘います。このような社会的闘争は、闘いの当事者同士が相手に致命的なダメージを与える前に、どちらかが降参して、当事者同士がお互いの優劣関係を受け入れたときに終息します。これまで、このような社会的闘争の終息がどのように制御されているのかは、全く分かっていませんでした。
例えば、2匹のオスのゼブラフィッシュを、24時間個別に飼った後、同じ水槽に移すと、両者は、一定の手順で体の大きさを強調する威示行動を行った後に、相手の側面へかみ付き攻撃を始めます。攻撃は数分で終了し、勝者と敗者が決まり、勝者は、水槽の中央の大部分を占拠して活発に泳ぎますが、敗者は、水槽の辺縁部でじっとするようになります(図1a)。一度勝敗が決まると、敗者は、新しい相手と対戦しても、大方負け続けるのに対して、勝者は、新しい相手に対しても勝ち続けるようになります。
手綱核は全ての脊椎動物で、間脳の最背側部に両側性で存在し、脳の情動に関わる神経核群(いわゆる大脳辺縁系の一部を含む)からの入力を受けて、中脳と後脳の境界部に接する腹側正中線上に一つだけ存在する脚間核や、さまざまなモノアミン神経細胞を含む神経核(背側被蓋野や縫線核)[3]への出力を中継します(図1b)。
岡本チームリーダーらはゼブラフィッシュを使った研究から、背側手綱核の外側と内側の二つの亜核は、それぞれが脚間核の背側と腹側に特異的に投射[4]して、外側亜核から背側脚間核への回路の強化は、降参せずに闘い続けるという勝者の行動選択を促進すること、内側亜核から腹側脚間核への経路の強化は、すぐに降参する敗者の行動選択を促進すること、さらにこの2本の回路がせめぎ合ってお互いを押さえようとする働きを持つことを明らかにしていました注1)。
手綱核から脚間核に至る2本の回路は、魚類だけでなく全ての脊椎動物で進化的に保存されて存在する、すなわち進化の過程で変わらずに保たれてきていることから、哺乳類の同種間の闘争行動も、この2本の回路によって制御されているのかが、次の課題となっていました(図1c、d)。
図1 ゼブラフィッシュとマウスでの手綱核-脚間核神経回路の保存
- (a)2匹のゼブラフィッシュによる社会的上下関係を巡る闘争の様式。
- (b)ゼブラフィッシュの脳での手綱核の各部(亜核)から脚間核への神経結合とその先への投射の様式。
- (c)ゼブラフィッシュとマウスの手綱核の進化的に相同な領域(亜核)の比較(同じ色の領域が相同)。
- (d)ゼブラフィッシュとマウスの手綱核-脚間核神経回路の形態的類似。上がゼブラフィッシュ、下がマウス。
- 注1)2016年4月1日プレスリリース「動物の争いでいつ降参するかを決める神経回路」
研究手法と成果
岡本チームリーダーらは、これまでの解剖学的研究から、ゼブラフィッシュの背側手綱核と腹側手綱核が、齧歯(げっし)類の手綱核の内側手綱核と外側手綱核に相当し、ゼブラフィッシュの背側手綱核の外側領域(亜核)dHbL(勝者の回路)がマウスなどの齧歯類の内側手綱核の背側領域dMHbと、ゼブラフィッシュの背側手綱核の内側領域dHbM(敗者の回路)が齧歯類の内側手綱核の腹側領域vMHbに相当することを明らかにしていました注2)(図1c)。
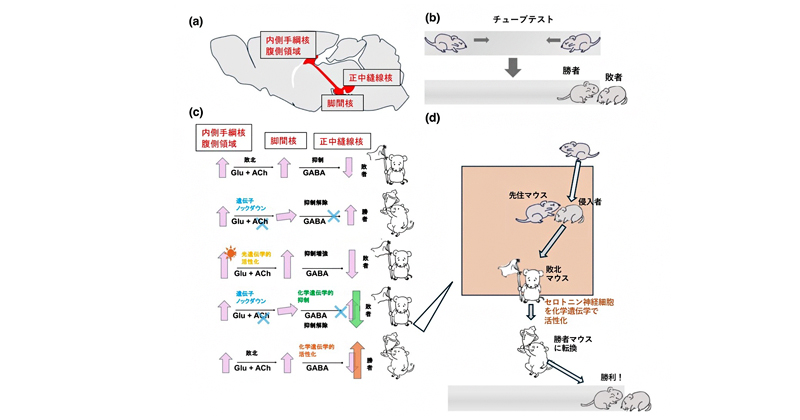
今回、共同研究グループは、これらの回路の機能的保存を検証するために、マウスの内側手綱核の腹側領域で、脚間核へ投射する神経の伝達物質として使われているアセチルコリンの産生を特異的に抑制した遺伝子改変マウス系統を作成し(図2a、c)、ゼブラフィッシュの敗者の回路と相同な回路を人為的に遮断することに成功しました。この系統は、チューブ内で相手を押し出すことによって勝敗を決めるチューブテスト(図2b)で、常に勝者となりました(図2c-ii、および動画注3)参照)。一方、光線の照射によってこの回路を人為的に活性化させる光遺伝学[5]という技術を使って、この回路を特異的に活性化すると、マウスは常にあっさり負けるようになりました(図2c-iii)。さらに、内側手綱核腹側領域のアセチルコリン産生を抑制したマウスでも、マウスでの敗者の回路の候補がさらにつながっている、正中縫線核のセロトニン神経細胞の活性を薬剤の投与によって特異的に抑制する化学遺伝学[6]という技術を適用すると、勝てなくなりました(図2c-iv)。
他のマウスがすんでいるケージに入れられて、先住のマウスから一方的に攻撃されて社会的敗北を喫したマウスは、その後チューブテストをすると常に敗北します。ところが、このようなマウスで、正中縫線核のセロトニン神経細胞を化学遺伝学的技術を用いて特異的活性化すると、勝つようになりました(図2c-v、dおよび動画注3)参照)。
図2 本研究の概要を表す模式図
- (a)マウスの脳で,内側手綱核、脚間核、正中縫線核のつながりを表す模式図。
- (b)チューブテストによるマウスの勝敗の決定。
- (c)本研究で行われた内側手綱核腹側領域への操作による、脚間核、正中縫線核の活性の変化と、勝敗への影響。Glu:グルタミン酸、ACh:アセチルコリン、GABA:ギャバ、抑制性の神経伝達物質。
- (d)一度敗北したマウスを勝者に変える方法。
これらの結果は、内側手綱核腹側領域から脚間核を経て正中縫線核に至る神経回路が、ゼブラフィッシュの敗者の回路と同じ働きを持つこと、敗北の状態に追い込まれたマウスも、この回路の人為的操作で、勝者として振る舞うように転換できることを示しています。
本研究によって、全ての動物で進化的に保存されている手綱核から脚間核に至る2本の回路が、全ての動物に共通して見られる社会的上下関係を巡る闘争の勝敗の制御において、機能的にも保存された重要な役割を果たしている可能性が明らかになりました。
- 注2)Amo, R., Aizawa, H., Takahoko, M., Kobayashi, M., Takahoko, R., Aoki, T., and Okamoto, H. (2010). Identification of the zebrafish ventral habenula as a homologue of the mammalian lateral habenula. J. Neurosci., 30:1566-1574.
この動画にはナレーションはありません
注3)動画 チューブテストによる勝敗の決定
前半は大きな野生型マウス(左、白)と小さな野生型マウス(右、茶)の闘い、後半は大きな野生型マウス(左、白)と敗者の回路を不活化した小さなマウス(右、茶)の闘い。
今後の期待
勝者の回路は、動物が諦めないで闘い続けられるように、動物のストレス耐性を高める働きがあると考えられます。一方、敗者の回路は、闘いをやめることを促進します。しかし敗者は、単に屈服や服従しているのではなく、危機的な状況を的確に判断し、自らはじっとしながら相手の動きを注意深く観察するように、行動様式を変化させています。すなわち、勝者の回路と敗者の回路は、相手と対峙(たいじ)する上で、攻撃モードと観察モードのどちらで対峙するかの選択に関わっていると考えられます。
手綱核から脚間核に至る2本の回路への入力や出力を行う神経回路がどのようなものかをさらに調べることによって、動物が、敵対的相手への対処の仕方をどのように決めているかを明らかにすることができます。また、この2本の回路が、どのようにせめぎ合い、どのようにして優位になる回路が決定されるのかを調べることによって、闘いを続けるかどうかの心の中の葛藤の仕組みも明らかにできると考えられます。
さらに、この2本の回路のバランスの崩れが、鬱やひきこもりなどの精神状態とどのように関わるのかを研究することによって、このような精神状態の脳科学的理解や治療法の開発に貢献すると期待されます。
補足説明
- 1.手綱核(たづなかく)-脚間核(きゃくかんかく)神経回路間脳にある手綱核から中脳にある脚間核へと投射する神経回路。マウスでは、手綱核の中でも特に内側手綱核が脚間核へ投射しており、情動の制御に関わる辺縁系からの入力を脳幹に伝達する経路と考えられている。脚間核はさらに、脳幹部のセロトニン系やアセチルコリン系を含むさまざまな領域へ投射を行っており、これらの情動制御と関わる脳領域の活動を調節していると考えられている。
- 2.正中縫線核(せいちゅうほうせんかく)のセロトニン神経細胞正中縫線核は、脳幹の正中部で脚間核の後方に位置する神経核である。セロトニン神経細胞を多く含み、大脳皮質、大脳基底核などさまざまな領域へ神経軸索を延ばして、セロトニンを送り出す。恐怖やストレスの制御に関与すると考えられている。
- 3.モノアミン神経細胞を含む神経核(背側被蓋野や縫線核)背側被蓋野脳幹部は、脚間核の背側に位置する広範な領域。ドーパミン、ノルアドレナリン、セロトニンなどのモノアミンに分類される神経伝達物質を持つ神経細胞群を含んでいる。
- 4.投射ある脳領域から別の脳領域に神経軸索を延ばしてつながること。
- 5.光遺伝学脳の特定の神経細胞に、光によって活性化されるタンパク分子を、遺伝学的手法を用いて発現させ、その神経細胞の興奮状態を光の照射によって操作する技術。
- 6.化学遺伝学脳の特定の神経細胞に人工的な受容体を発現させ、その受容体を特異的に刺激する薬物を全身に投与することで、特定の神経細胞の活動を人工的に操作する技術。
共同研究グループ
理化学研究所 脳神経科学研究センター 意思決定回路動態研究チーム(研究当時) チームリーダー(研究当時)岡本 仁(オカモト・ヒトシ) (現 知覚運動統合機構研究チーム 客員主管研究員、理研 名誉研究員、早稲田大学 理工学術院 客員上級研究員、研究院客員教授、公益財団法人 神経研究所 客員研究員) 研究員(研究当時)松股 美穂(マツマタ・ミホ) (現 広島大学 大学院医系科学研究科 神経生物学研究室 助教) 研究員(研究当時)平尾 顕三(ヒラオ・ケンゾウ) (現 ラクオリア創薬株式会社 研究員) 研究員(研究当時)小林 琢磨(コバヤシ・タクマ) (現 武田薬品工業株式会社 研究員) 大学院生リサーチ・アソシエイト(研究当時)半田 剛久(ハンダ・タケヒサ) (現 東京科学大学 精神科(大学院 精神行動医科学分野)特任助教) 研究員(研究当時)杉山 拓(スギヤマ・タク) (現 理研 脳神経科学研究センター 生体物質分析支援ユニット 専門技術員) 研究員(研究当時)柿沼 久哉(カキヌマ・ヒサヤ) (現 理研 脳神経科学研究センター 知覚運動統合機構研究チーム テクニカルスタッフⅠ) テクニカルスタッフⅠ(研究当時)タンビル・イスラム(Tanvir Islam) (現 理研 脳神経科学研究センター 生体物質分析支援ユニット テクニカルスタッフⅠ) 行動遺伝学技術開発チーム(研究当時) 研究員(研究当時)小林 祐樹(コバヤシ・ユウキ) (現 理研 脳神経科学研究センター センター長室 高度研究支援専門職) 神経回路・行動生理学研究チーム テクニカルスタッフⅠ アーサー・ファン(Arthur J. Huang) 基礎科学特別研究員 イイン・シュウ(Yijun Zhou)
チームディレクター トーマス・マックヒュー(Thomas J. McHugh)
関連記事:


コーヒー成分が老化や病気に関わる受容体に作用する可能性

